


This past May, I was fortunate enough to take part in a conference in Venice, Italy, which was a retrospective on the legacy of famous paleontologist and author Stephen Jay Gould 10 years after his death. The choice of Venice as the conference venue was a nod to one of Gould’s most famous and influential papers, “The Spandrels of San Marco and the Panglossian Paradigm: A Critique of the Adaptationist Programme“, which he co-authored with Richard Lewontin in 1979.
In the paper, Gould and Lewontin (1979) drew an analogy between the “spandrels” (or, if you’re overly pedantic about architectural terminology, “pendentives“) in Saint Mark’s Basilica in Venice and features of organisms that are often assumed to represent adaptations whose current functions reflect the reason for their origin. The spandrels in St. Mark’s Basilica house elaborate mosaics depicting important Christian iconography, and represent one of the most stunning aspects of the cathedral.
However, housing religious imagery is not why the spandrels were created in the first place. As Gould and Lewontin (1979) point out, spandrels are the inevitable result of resting a domed ceiling atop walls with arched doorways. Once they exist, spandrels can be co-opted for a function such as artistic decoration — but whatever this current function may be, it does not, by itself, explain the origin of the structures.
Many traits of organisms, rather than being the result of adaptive evolution under selection for their current function (if any) are, in fact, biological “spandrels”.

Yours truly enjoying the amazing artistry in St. Mark’s Basilica.
The tendency to conflate current function with historical origin is one of the things that Gould and Lewontin saw as a pervasive problem in the evolutionary biology of the 1970s. They also considered the general tendency to focus almost exclusively on adaptationist explanations for individual traits to be misguided.
Not only did authors not properly consider alternative, non-adaptive explanations, they presented only weak evidence in support of adaptive “just-so stories” or simply moved on to a new adaptive tale if evidence was found lacking for a particular explanation.
Abstract
An adaptationist programme has dominated evolutionary thought in England and the United States during the past 40 years. It is based on faith in the power of natural selection as an optimizing agent. It proceeds by breaking an organism into unitary ‘traits’ and proposing an adaptive story for each considered separately.
Trade-offs among competing selective demands exert the only brake upon perfection; non-optimality is thereby rendered as a result of adaptation as well. We criticize this approach and attempt to reassert a competing notion (long popular in continental Europe) that organisms must be analysed as integrated wholes, with Bauplane so constrained by phyletic heritage, pathways of development and general architecture that the constraints themselves become more interesting and more important in delimiting pathways of change than the selective force that may mediate change when it occurs.
We fault the adaptationist programme for its failure to distinguish current utility from reasons for origin (male tyrannosaurs may have used their diminutive front legs to titillate female partners, but this will not explain why they got so small); for its unwillingness to consider alternatives to adaptive stories; for its reliance upon plausibility alone as a criterion for accepting speculative tales; and for its failure to consider adequately such competing themes as random fixation of alleles, production of non-adaptive structures by developmental correlation with selected features (allometry, pleiotropy, material compensation, mechanically forced correlation), the separability of adaptation and selection, multiple adaptive peaks, and current utility as an epiphenomenon of non-adaptive structures.
We support Darwin’s own pluralistic approach to identifying the agents of evolutionary change.
Some subsequent authors have charged that Gould and Lewontin (1979) presented a straw man, and did not provide a fair assessment of what members of the so-called adaptationist camp actually did or said. Nevertheless, I think many biologists feel that theirs was a very important contribution, at least as a warning for how the study of trait evolution could go awry. I would take a stronger position, and argue that not only was the Gould and Lewontin (1979) paper needed at the time, but that it is still needed today.
And nowhere is this more true than in studies of human behavior and anatomy, particularly because these tend to be widely covered in the science media. Evolutionary psychology takes a large amount of flak from evolutionary biologists and others for its perceived tendency to present “just-so stories” or to make extraordinary assumptions about the social and physical habitats of hominin ancestors. But this is not limited to studies of human minds by any means.
I recently discussed one example, the widely-reported claim that the dimensions of human hands evolved in large part to allow the formation of fists. In that post, I outlined several major problems with the underlying assumptions and the (very weak) data that were presented as supporting evidence. I won’t re-hash these points here, but many of them apply equally to the even more recent claim that a human trait is the product of adaptive evolution: namely that the wrinkling of our fingers and toes when submerged in water is an adaptation for gripping in wet conditions.
The idea was first put forward by Mark Changizi and colleagues of 2AI Labs in Boise, Idaho. Changizi has a PhD in applied math and primarily studies neurobiology, but in this case his focus was on why fingers wrinkle when they’re soaked. Specifically, Changizi et al. (2011) wondered whether the wrinkled patterns that form on the fingers after prolonged soaking have a function similar to the treads on tires, namely for channelling water and improving grip in wet conditions.
Based on the idea that wrinkled digits might actually represent water channels, they looked at photos of 28 wrinkly fingers from 13 hands that they found online. They then compared these wrinkle patterns to the channels of rivers in a mountainous drainage system and to the treads on car tires and found that they are similar in that they branch and diverge from the middle of the finger.
.")
From Changizi et al. (2011).
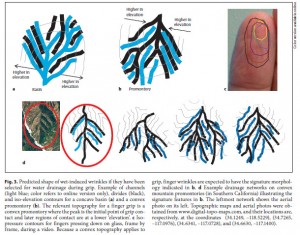
From Changizi et al. (2011).
In addition, the authors noted that the wrinkling of fingers is not simply a result of osmotic changes when hands are soaked, and that there is evidence that the process is linked to the action of the autonomic nervous system. Or, at least, wrinkling does not occur if there is nerve damage in the hand. In fact, physicians use wrinkling in water as a test of neural function in the limbs (Wilder-Smith 2004; Barneveld et al. 2010). So, the evidence to this point was a) wrinkling seems to be under nervous control, and b) the wrinkles look like drainage systems or tire treads.
This hypothesis has gained more attention recently with the publication of a follow up study by Kareklas et al. (2013) in the lab of Tom Smulders in the Institute of Neuroscience at Newcastle University. Kareklas et al. (2013) wanted to test the idea that wrinkling improves grip in wet conditions. To do so, they had 20 volunteers soak their hands in 40°C water for 30 minutes and then had them move marbles of different sizes from one container to another, with the source container either filled with water or dry. They compared this to the performance of the volunteers in the same task when their hands had not been soaked and therefore their fingers were not wrinkled.
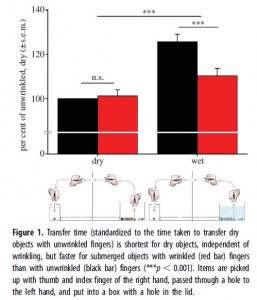
From Kareklas et al. (2013).
Here is the entire Results section of the paper:
It took participants anywhere between 72 s (fastest person) and 198 s (slowest person) to transfer all the items. All participants transferred dry objects more quickly than submerged objects, taking on average 17 per cent (±2.3% s.e.m.) more time to transfer submerged objects (F1,16 = 35.80, p < 0.001).
With wrinkled fingers, transfer of submerged objects happened in 12 per cent (±2.3% s.e.m.) less time than with unwrinkled fingers. There was no difference in the time it took to transfer dry objects with or without wrinkles on the fingers (interaction: F1,16 = 44.10, p < 0.001; figure 1).
This finding shows a clear advantage of having wrinkled fingers when manipulating submerged objects, but not dry objects.
In short, soaking your hands in 40°C water for 30 minutes will give you a 12% advantage in picking up round, smooth objects if they are underwater.
Not surprisingly, given that it is about a familiar yet curious human trait, the science media have reported this study widely.
Now, as you may have guessed, I don’t find this hypothesis compelling at all, for many reasons. The first is that it is based on the thinnest of datasets. Photos of 13 hands with finger wrinkles that resemble drainage channels. And a small advantage in manipulating wet marbles. That’s it for the data.
I also don’t buy the conceptual arguments, for example Changizi’s claim that there are infinitely many possible patterns for wrinkles to form, and yet we see the one that is consistent with rain treads. I strongly doubt that there are more than a few possible arrangements for wrinkles, given the properties of human skin and the shape of human digits. Moreover, it looks to me like there is substantial variability in wrinkle patterns even within the tiny sample of their study.
Would all of these channel water effectively? We don’t know, because no effort was taken to demonstrate that wrinkles actually displace water. (I have my doubts about this. Treads are hard, fingers are soft. Tires rotate, fingers don’t).
Finally, I am not convinced that the link to the central nervous system implies that this trait is functional. Vasoconstriction is involved (Wilder-Smith and Chow 2003a,b), and this can be controlled by the nervous system, but that alone does not mean that the trait evolved as an adaptation rather than as a spandrel. Perhaps the effect is simply a byproduct of other nerve signal changes that are functional in response to hot water immersion.
I don’t know, but neither does anyone else because such non-adaptive hypotheses have not been considered. There certainly seems to be some cross-wiring going on, since reduced wrinkling ability is correlated with congestive heart failure (Kamran et al. 2011) and heart rate variability (Win et al. 2010).
There is also this:
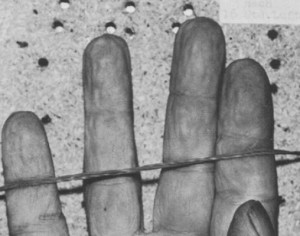
From Reh (1984).
The photo above shows a pretty typical wrinkle pattern for a hand that has been submerged in water (in this case, for 120 minutes in 13°C water). What sets this hand apart from the ones examined by Changizi et al. (2011) is that it belongs to a dead person. That is to say, wrinkling occurs, with the same pattern and at a similar rate, even when the subject is deceased (Weber and Laufkötter 1984; Reh 1984).
From Reh (1984):
Abstract
Studies were carried out systematically in both corpses and the hands of corpses in order to find out when ” washerwoman ‘s skin” begins. The temperatures ranged between 10 degrees and 18 degrees C and the time of the experiments did not exceed 300 min. The initial formation of washerwoman ‘s skin could be observed after 20-30 min at the fingertips and after 50-60 min in the entire finger. The longest intervals observed were 100 or 150 min, respectively.
These long intervals were interpreted as being exceptions from the norm, probably due to either extraordinarily heavy strips of fat on the fingers or abnormally hard skin on the hands. The course of washerwoman‘s skin obviously depends on the water temperature, and afterwards it disappears rather slowly–even after a short immersion–in the open air.
We will carry out additional experiments in the future.
If finger and toe wrinkling is an adaptation for gripping in the wet as claimed, we might also ask:
- Why does it take so long to occur? Wouldn’t it be better to have a quick response rather than a system that takes at least a few minutes of immersion to kick in? (I stood in the warm shower for 10 minutes this morning, and my feet did not wrinkle at all.)
- Why is the response strongest in very warm water (Cales et al. 1997)? Is 40°C water really encountered a lot under natural conditions of foraging or running?
- If this is an adaptation for simply channelling water, why is the response strongly affected by the salinity and pH of the water (Tsai and Kirkham 2005)?
I don’t think these observations provide support for the idea that finger wrinkling is an adaptive response to being wet, or at least they are not what I would expect to be the case if this were such an adaptation.
In any case, the bigger problem with the wrinkled gripper ape hypothesis, the punching ape hypothesis, and other such hypotheses is that they are not based on solid evolutionary thinking. More specifically, the authors of these hypotheses do not seem to work through the assumptions that would have to be true for these traits to be adaptive in the way that they propose.
Did an ancestral population actually have variation in this trait, and is it heritable? Did these ancestors really submerge their hands and feet regularly? Did individuals with more wrinkles actually have more children than individuals with less tread-like wrinkles?
Many biologists favour the adaptationist approach to studying trait evolution. That is, they begin with ideas about why a certain trait might be adaptive and then proceed from there. Others (increasingly, myself included) may argue that there must be evidence that the trait is adaptive in the first place before such hypotheses are pursued.
This may be a matter of preference or training or philosophical bent more than anything. However, it should be clear either way that there are good and bad ways to test adaptationist hypotheses.
Here’s the bad way:
- Notice a trait and think up an adaptive function for it.
- Assemble a small number of modern humans and get them to do whatever it is you think the trait is functional for (say, punching a heavy bag or picking up wet marbles). Make this unrealistic in terms of ancestral environments (e.g., use trained martial artists for punching or soak in 40°C water and handle glass marbles) and don’t control for other factors (say, warming up the hand).
- Compare this as a binary trait rather than considering variation. For example, a buttressed fist vs. an open palm or fully wrinkled fingers vs. dry fingers.
- Find a very weak effect, even with this extreme comparison.
- Conclude that the trait in question evolved for the hypothesized function and ignore the obvious assumptions that are required for such a conclusion to hold.
Here’s a better way:
- Notice a trait and determine the extent to which it varies within the study species.
- Study the relationship between variation in the trait and performance of the proposed function. This would have been very easy for the marble study: soak hands for a few minutes to get partial wrinkles and assess performance, then soak for longer, etc. If this trait evolved as an adaptation, then differences in wrinkliness within the same finger should correlate with grippiness of that finger. (Here’s an example of that approach in studying the beaks of crossbills)
- Explicitly address the necessary assumptions about ancestral traits, habitat, selective coefficients, and population size.
- Use the comparative method within a phylogenetic context. Identify other species that have the trait and those that lack it. Has the trait evolved independently under similar conditions? Is it found in species that would not be expected to exhibit it if it evolves adaptively for a particular function?
- Consider and rule out non-adaptive explanations (developmental constraints, pleiotropy, etc.) as much as possible.
Getting the data for this second list is much more difficult than for the first, but without them any conclusions about adaptive evolutionary history remain subject to the criticisms laid out several decades ago by Gould and Lewontin (1979).
References
Cales, L., R.A. Weber, and T.X. Temple (1997). Effects of water temperature on skin wrinkling. J. Hand Surg. 22A: 747-749.
Changizi, M., R. Weber, R. Kotecha, and J. Palazzo (2011). Are wet-induced wrinkled fingers primate rain treads? Brain Behav. Evol. 77: 286-290.
Gould, S.J. and R.C. Lewontin (1979). The Spandrels of San Marco and the Panglossian Paradigm: A Critique of the Adaptationist Programme. Proc. R. Soc. Lond. B 205: 581-598.
Kamran, H., L. Salciccioli, and J.M. Lazar (2011). Reduced water induced skin wrinkling in congestive heart failure. Clin. Auton. Res. 21: 361-362.
Kareklas, K., D. Nettle, and T.V. Smulders (2013). Water-induced finger wrinkles improve handling of wet objects. Biol. Lett., in press.
Reh, H. (1984). On the early postmortal course of “washerwoman’s skin” at the fingertips [in German]. Z. Rechtsmed. 92: 183-188.
Tsai, N. and S. Kirkham (2005). Fingertip skin wrinkling — the effect of varying tonicity. J. Hand Surg. 30B: 273-275.
Van Barneveld, S., J. van der Palen, and M.J.A.M. van Putten (2010). Evaluation of the finger wrinkling test: a pilot study. Clin. Auton. Res. 20: 249-253.
Weber, W. and R. Laufkötter (1984). Stage classification of washerwoman’s hands at experimental time intervals [in German]. Z. Rechtsmed. 92: 277-290.
Wilder-Smith, E.P.V. and A. Chow (2003a). Water-immersion wrinkling is due to vasoconstriction. Muscle Nerve 27: 307-311.
Wilder-Smith, E. and A. Chow (2003b). Water immersion and EMLA cause similar digit skin wrinkling and vasoconstriction. Microvascular Res. 66: 68-72.
Wilder-Smith, E.P.V. (2004). Water immersion wrinkling: Physiology and use as an indicator of sympathetic function. Clin. Auton. Res. 14: 125-131.
Win, S., L. Salciccioli, H. Kamran, P. Baweja, M. Stewart, and J.M. Lazar (2010). Water immersion-induced skin wrinkling is related to heart rate variability. Cardiology 116: 247-250.
Originally appeared on EvolverZone January 11, 2013



Comments