Our energy dilemma
The hydrocarbon economy is faltering as oil reserves dwindle worldwide (Hirsch, 2008). Commodity prices have begun to fluctuate drastically due to the uncertain cost of petroleum, which resulted in food riots around the world in 2008. With a steadily decreasing energy supply and the demands on energy systems continually growing, the planet is in dire economic, geopolitical, and environmental straits. In order to halt the advance of climate change, prevent ecological collapse, rescue the global economy, and ensure our energy security, humanity must find a way to harness currently available (non-fossilized) energy. The largest source of energy on Earth, excluding the future potential for thermonuclear fusion reactors, is the sun. Human civilization consumes 15 TW annually while approximately 80,000 TW of solar energy fall on the Earth’s surface each year (Makarieva et al., 2008). For hundreds of millions of years, this solar flux has been the driving force for life on Earth. It is estimated that photosynthesis absorbs and distributes seven times more energy to the biosphere (~100 TW) than is consumed anthropogenically each year (Makarieva et al., 2008). There are many promising technologies in development that will cheaply harness solar output, like next generation photovoltaics, advanced wind turbines, tidal power systems, wave energy generators, solar thermal collectors, biofuels, solar fuels, electrofuels, etc. In the end, some combination of these solutions, coupled with improved energy and transportation infrastructure, will be needed to resolve our energy dilemma. In this article, I focus on cyanobacteria and their potential for hydrogen production, which, if properly developed, could provide sustainable fuel for transportation and industry in a post-petroleum world.

BioHydrogen Airship of the Future?
The subjects of this Earth-transforming story, and the only organisms known to have evolved oxygenic photosynthesis, are the cyanobacteria (Nisbet et al., 2007). For a billion years, the electrons in water sat untouched by photosynthetic life because it was energetically unfeasible to oxidize water and reduce NADP+ (an electron carrier needed for the reduction of inorganic carbon) in a single step. In an evolutionary leap, cyanobacteria pioneered the coupling of two photosynthetic reaction centers, P680 and P700 (named for the wavelengths of light they optimally absorb), referred to as photosystem II (PSII) and photosystem I (PSI), respectively (Allen and Martin, 2007; Mimuro et al., 2008). Thus, the redox problem was resolved by first pumping low-energy electrons up the redox gradient to a temporary reservoir (the quinone pool/electron transfer chain), from which the second photosystem (PSI) could pull out excited electrons in order to transfer them to an even higher energy state, sufficient for the reduction of NADP+ to NADPH + H+.
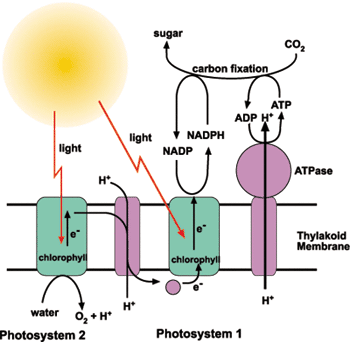
The rising concentration of oxygen in the atmosphere presented new challenges to early life on Earth. Most species at the time were obligate anaerobes or microaerophiles, and only those organisms that had the ability to deal with highly reactive oxygen radicals and persist in an oxygen-enriched atmosphere would inherit the majority of Earth’s habitats (Brioukhanov and Netrusov, 2007; Bendall et al., 2008). Cyanobacteria, in addition to overcoming the general toxicity of oxygen (a byproduct of their new metabolism), had to find a way to acquire nitrogen under aerobic conditions (fixing atmospheric nitrogen gas into biologically-available ammonia). The molybdenum-iron nitrogenase enzyme is highly conserved among diazotrophs (nitrogen-fixers) and is very sensitive to oxygen, which destroys the activity of the enzyme. Cyanobacteria got around this issue by separating oxygenic photosynthesis and nitrogen fixation, either spatially or temporally (Tsygankov, 2007). Some cyanobacteria only activate nitrogen fixation under dark anaerobic conditions, when PSII is unable to evolve oxygen. Other cyanobacteria, like filamentous Nostoc punctiforme, form a specialized cell type for nitrogen fixation, called a heterocyst. Heterocysts deactivate their PSII complexes, grow thickened cell walls, and exhibit higher intracellular respiration rates, which keep oxygen levels very low (Cardona, 2009). Vegetative cells provide the heterocysts with carbohydrates, while the heterocysts provide the vegetative cells with fixed nitrogen (Cardona, 2009).
Nostoc punctiforme
Nitrogenase is responsible for the following reaction: N2 + 8 H+ + 8 e− + 16 ATP → 2 NH3 + H2 + 16 ADP + 16 Pi (Tikhonovich and Provorov, 2007). This process is very energy-intensive, requiring 8 moles of ATP for every mole of ammonia produced. In addition to ammonia, the nitrogenase enzyme also produces a molecule of hydrogen gas for every molecule of gaseous nitrogen that it fixes. Hydrogen production via nitrogenase is relatively inefficient, due to the large amounts of ATP required, but it can still be used to produce measurable amounts molecular hydrogen. One impediment to producing hydrogen in this way is the presence of uptake-hydrogenases (Schütz et al., 2004), which oxidize nitrogenase-produced hydrogen in order to minimize the energy loss from N-fixation by reclaiming ATP via the oxyhydrogen reaction, removing oxygen from the interior of the cell and providing reducing equivalents for other cellular processes (Tamagnini et al., 2007). Concordantly, uptake-hydrogenase enzyme activity has been shown to correlate with nitrogenase activity (Schütz et al., 2004). Therefore, it is not surprising that a Nostoc uptake-hydrogenase knockout mutant exhibits a significantly higher hydrogen output than the wild-type (Lindberg et al., 2004).
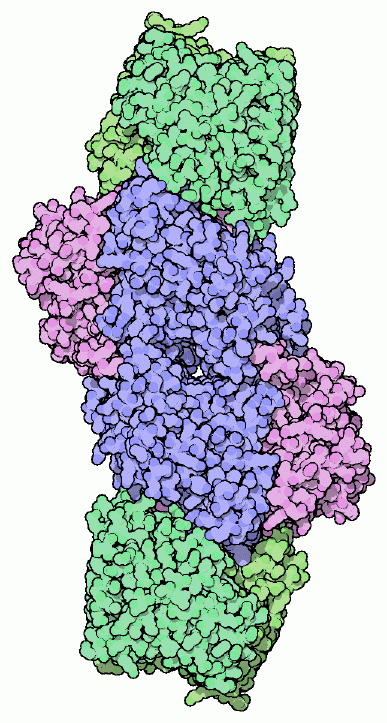
Nitrogenase
Nostoc punctiforme was isolated from a mutualistic association with a cycad (Costa and Lindblad, 2002). Nostoc, when living symbiotically, has a higher heterocyst frequency than it does as a free-living organism, and will devote much of its metabolism to the production and secretion of specific nitrogen-rich metabolites that are beneficial to its plant host (Enderlin and Meeks, 1983). Since Nostoc has evolved to mass-produce a specific metabolite when living symbiotically, it seems almost pre-programmed for fuel production. Heterocystous N-fixing cyanobacteria have minimal nutritional requirements, high photosynthetic efficiencies and can create anoxic microenvironments inside specialized cells that allow anaerobic processes to occur under aerobic conditions. All of these attributes are conducive to affordable bioreactor design for hydrogen production. If Nostoc hydrogen metabolism can be effectively engineered, using novel synthetic biology tools, the door to developing successful photobiological hydrogen-yielding technologies is opened. 
Nostoc living symbiotically in plant tissue.
An astounding result of recent genomic sequencing projects is that the length of a genome does not predict the morphological or physiological complexity of an organism. For example, length of the human genome is similar to that of the fruit fly (Mukherji and van Oudenaarden, 2009). Instead, it has been found that biological modularity can help explain the diversity of form and function in the natural world (Mukherji and van Oudenaarden, 2009). A limited subset of predictable biological “parts” can be assembled in various ways to produce molecular “devices”, which can be arranged into “systems” to carry out different functions. In this light, one can view a living cell as a combination of co-regulated genetic circuits, working in tandem. Another surprising discovery made recently shows the inherent resiliency of biological circuitry to rewiring. Isalan et al. (2008), in order to test the limits of perturbing regulatory networks in biological systems, found that randomly rewiring Escherichia coli transcriptional networks by synthetically altering transcription factor/promoter pairings only resulted in faulty growth in 5% of cases. This level of tolerance to random restructuring of biological networks is highly conducive to the adaptation and evolvability of living systems by allowing large-scale alterations to be made to an organisms genome without significantly impeding its growth (Isalan et al., 2008; Mukherji and van Oudenaarden, 2009). Biological modularity and network resiliency provide powerful mechanisms for the rapid evolution of novel or optimized processes by reshuffling pre-existing genes/proteins (Isalan et al., 2008; Mukherji and van Oudenaarden, 2009; Picataggio, 2009). 
Transcriptase enzymes moving along the DNA strand, from a recent synthetic biology comic book published in Nature.
Synthetic biologists have already succeeded in characterizing thousands of biological parts with defined functions and performance parameters, which can be accessed openly at the wiki site http://partsregistry.org/ maintained by the Massachusetts Institute of Technology, but they are not yet capable of engineering whole biological systems with the same precision and reliability that, say, electrical engineers are accustomed to. One of the main challenges is to identify the subset of genes that are absolutely necessary for the survival of a minimal genome - the smallest number of genes that allows for the replication of an organism in a particular environment (Cho et al., 1999). Until researchers can build a living cell from the ground up, they won’t totally understand the limits of metabolic engineering. The immense potential for engineering crucial synthetic metabolic circuits has already been demonstrated by the work of Dr. Jay Keasling, who produced an important precursor to the anti-malarial drug artemisinin in E. coli (Hale et al., 2007). Keasling’s work has cut the cost of artemisinin by ten fold and will provide many people in the third world with access to crucial malaria treatments, literally saving millions of lives. 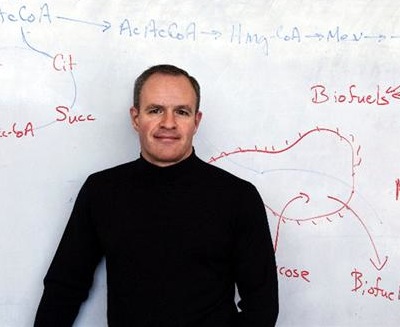
Dr. Jay Keasling
The creation of standard biological parts (promoters, ribosomal binding sites, repressors, activators, etc.) for cyanobacteria is simplified by the work already completed in E. coli and by the current open-source nature of synthetic biology resources, fostered by organizations like the BioBricks Foundation (http://bbf.openwetware.org/) and iGEM (http://2009.igem.org). With our current understanding of transcriptional and translational regulation in cyanobacteria, we can re-design genetic regulatory elements and codon-optimised genes from distantly related organisms in silico and synthesize these constructs in vitro for expression in cyanobacteria. A team in France has developed codon-optimized (for expression in E. coli, Synechocystis, Nostoc and Anabaena) synthetic [Fe-Fe] hydrogenase genes from Chlamydomonas reinhardtii and Clostridium acetobutylicum that have been linked to a synthetic ferredoxin ligands derived from a chlamydomonal ferredoxin (Jaramillo, A., 2009, École Polytechnique, Palaiseau, France, privileged information). Recently, researchers in the Department of Photochemistry and Molecular Science at Uppsala University have characterized Ptrc promoters, derived from the lacUV5 promoter, ribosomal binding sites and an expression vectors (pPMQAK1 and pPMQAC1) that are broadly functional in E. coli and in cyanobacteria (Brosius et al., 1985; Huang et al., 2010; Gibbons, 2010). With these tools, it is now possible to express prolific [Fe-Fe] hydrogenases and their respective maturation proteins in cyanobacteria. It is only a matter of time before viable hydrogen-yielding systems are in place. 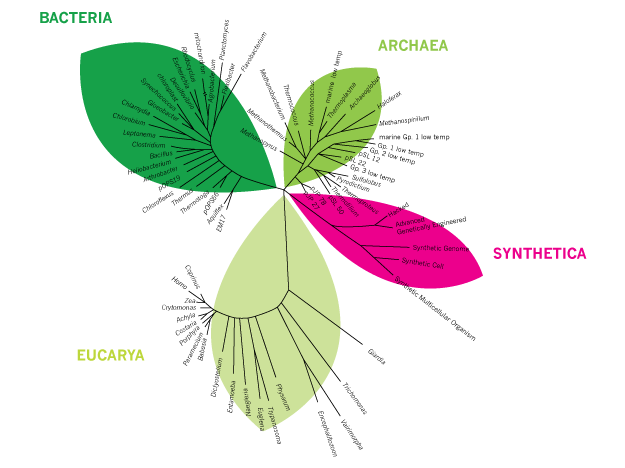
Synthetica, a novel branch to the tree of life.
In this post, I wanted to provide a glimpse of recent developments in photobiological hydrogen production (a field that I'm familiar with), but this is just one small piece of the puzzle. There are many teams of brilliant researchers around the globe chipping away at their own corners of sustainable energy. The future green economy will have to take a multi-faceted approach to energy, incorporating dozens of the most successful technologies, alongside behavioral and policy reform (i.e. less consumption, governmental regulation of greenhouse gasses, more localized food production, ecosystem preservation/restoration etc.). My point was to inspire hope, despite all the doom and gloom surrounding climate change, by illuminating a subset of the work and ideas of our best and brightest, who toil day and night to build a better tomorrow. While we cannot ignore the perils that lie ahead, we must maintain a certain degree of hope in order to keep our heads above water and look towards a better horizon. Fear not, fellow humans - for every Senator James Inhofe, there are a hundred thousand scientists, social activists, community horticulturists and conservationists. By hook or by crook, we will shape our world into a more verdant and peaceful place (or die trying). We've got soul, but we're not soldiers.
Citations
Agapakis, C., Ducat, D., Boyle, P., Wintermute, E., Way, J.,&Silver, P. (2010). Insulation of a synthetic hydrogen metabolism circuit in bacteria Journal of Biological Engineering, 4 (1) DOI: 10.1186/1754-1611-4-3
Allen, J.F., and Martin, W. (2007) Out of thin air. Nature 445: 630-632.
Asada, Y. (2000). Heterologous expression of clostridial hydrogenase in the cyanobacterium Synechococcus PCC7942 Biochimica et Biophysica Acta (BBA) - Gene Structure and Expression, 1490 (3), 269-278 DOI: 10.1016/S0167-4781(00)00010-5
Atsumi, S., Higashide, W.,&Liao, J. (2009). Direct photosynthetic recycling of carbon dioxide to isobutyraldehyde Nature Biotechnology, 27 (12), 1177-1180 DOI: 10.1038/nbt.1586
Bendall, D., Howe, C., Nisbet, E.,&Nisbet, R. (2008). Introduction. Photosynthetic and atmospheric evolution Philosophical Transactions of the Royal Society B: Biological Sciences, 363 (1504), 2625-2628 DOI: 10.1098/rstb.2008.0058
Blankenship, R.,&Hartman, H. (1998). The origin and evolution of oxygenic photosynthesis Trends in Biochemical Sciences, 23 (3), 94-97 DOI: 10.1016/S0968-0004(98)01186-4
Brioukhanov, A.,&Netrusov, A. (2007). Aerotolerance of strictly anaerobic microorganisms and factors of defense against oxidative stress: A review Applied Biochemistry and Microbiology, 43 (6), 567-582 DOI: 10.1134/S0003683807060014
Brosius, J., Erfle, M., and Storella, S. (1985) Spacing of the -10 and -35 regions in the tac promoter: effect on its in vivo activity. The Journal of Biological Chemistry 260: 3539-3541.
Cardona, T. (2009) The heterocysts of Nostoc punctiforme: from proteomics to energy transfer. Doctoral Dissertation, Uppsala University.
Cho, M. (1999). GENETICS:Ethical Considerations in Synthesizing a Minimal Genome Science, 286 (5447), 2087-2090 DOI: 10.1126/science.286.5447.2087
Cleaves, H., Chalmers, J., Lazcano, A., Miller, S., & Bada, J. (2008). A Reassessment of Prebiotic Organic Synthesis in Neutral Planetary Atmospheres Origins of Life and Evolution of Biospheres, 38 (2), 105-115 DOI: 10.1007/s11084-007-9120-3
Cohen, M., Wallis, J., Campbell, E., & Meeks, J. (1994). Transposon mutagenesis of Nostoc sp. strain ATCC 29133, a filamentous cyanobacterium with multiple cellular differentiation alternatives Microbiology, 140 (12), 3233-3240 DOI: 10.1099/13500872-140-12-3233
Costa, J.-L., and Lindblad, P. (2002) Cyanobacteria in Symbiosis with Cycads. In Cyanobacteria in Symbiosis, pp. 195-205. Netherlands: Springer
Enderlin, C., & Meeks, J. (1983). Pure culture and reconstitution of the Anthoceros-Nostoc symbiotic association Planta, 158 (2), 157-165 DOI: 10.1007/BF00397709
Fontecilla-Camps, J., Amara, P., Cavazza, C., Nicolet, Y., & Volbeda, A. (2009). Structure–function relationships of anaerobic gas-processing metalloenzymes Nature, 460 (7257), 814-822 DOI: 10.1038/nature08299
Ghirardi, M., Posewitz, M., Maness, P., Dubini, A., Yu, J., & Seibert, M. (2007). Hydrogenases and Hydrogen Photoproduction in Oxygenic Photosynthetic Organisms
Annual Review of Plant Biology, 58 (1), 71-91 DOI: 10.1146/annurev.arplant.58.032806.103848
Gibbons, S.M. (2010) Synthetic biology for renewable energy: engineering hydrogen production in cyanobacteria. Masters Thesis. Advisors: Lindblad, P., Heidorn, T. Uppsala University, Sweden.
Hale, V., Keasling, J.D., Renninger, N., and Diagana, T.T. (2007) Microbially Derived Artemisinin: A Biotechnology Solution to the Global Problem of Access to Affordable Antimalarial Drugs. Am J Trop Med Hyg 77: 198-202.
Hirsch, R. (2008). Mitigation of maximum world oil production: Shortage scenarios Energy Policy, 36 (2), 881-889 DOI: 10.1016/j.enpol.2007.11.009
Huang, H., Camsund, D., Lindblad, P., & Heidorn, T. (2010). Design and characterization of molecular tools for a Synthetic Biology approach towards developing cyanobacterial biotechnology Nucleic Acids Research, 38 (8), 2577-2593 DOI: 10.1093/nar/gkq164
Isalan, M., Lemerle, C., Michalodimitrakis, K., Horn, C., Beltrao, P., Raineri, E., Garriga-Canut, M., & Serrano, L. (2008). Evolvability and hierarchy in rewired bacterial gene networks Nature, 452 (7189), 840-845 DOI: 10.1038/nature06847
King, P., Posewitz, M., Ghirardi, M., & Seibert, M. (2006). Functional Studies of [FeFe] Hydrogenase Maturation in an Escherichia coli Biosynthetic System Journal of Bacteriology, 188 (6), 2163-2172 DOI: 10.1128/JB.188.6.2163-2172.2006
Lindberg, P., Lindblad, P., and Cournac, L. (2004) Gas Exchange in the Filamentous Cyanobacterium Nostoc punctiforme Strain ATCC 29133 and Its Hydrogenase-Deficient Mutant Strain NHM5. Appl Environ Microbiol 70: 2137-2145.
MAKARIEVA, A., GORSHKOV, V., & LI, B. (2008). Energy budget of the biosphere and civilization: Rethinking environmental security of global renewable and non-renewable resources Ecological Complexity, 5 (4), 281-288 DOI: 10.1016/j.ecocom.2008.05.005
Mimuro, M., Tomo, T., & Tsuchiya, T. (2008). Two unique cyanobacteria lead to a traceable approach of the first appearance of oxygenic photosynthesis Photosynthesis Research, 97 (2), 167-176 DOI: 10.1007/s11120-008-9311-4
Mukherji, S., & van Oudenaarden, A. (2009). Synthetic biology: understanding biological design from synthetic circuits Nature Reviews Genetics DOI: 10.1038/nrg2697
NISBET, E., GRASSINEAU, N., HOWE, C., ABELL, P., REGELOUS, M., & NISBET, R. (2007). The age of Rubisco: the evolution of oxygenic photosynthesis Geobiology, 5 (4), 311-335 DOI: 10.1111/j.1472-4669.2007.00127.x
Oliveira, P., & Lindblad, P. (2007). An AbrB-Like Protein Regulates the Expression of the Bidirectional Hydrogenase in Synechocystis sp. Strain PCC 6803 Journal of Bacteriology, 190 (3), 1011-1019 DOI: 10.1128/JB.01605-07
Picataggio, S. (2009). Potential impact of synthetic biology on the development of microbial systems for the production of renewable fuels and chemicals Current Opinion in Biotechnology, 20 (3), 325-329 DOI: 10.1016/j.copbio.2009.04.003
Pierson, B.K., and Olson, J.M. (1989) Evolution of photosynthesis in anoxygenic photosynthetic prokaryotes. Washington, D.C.: American Society for Microbiology.
Schutz, K., Happe, T., Troshina, O., Lindblad, P., Leitao, E., Oliveira, P., & Tamagnini, P. (2004). Cyanobacterial H2 production ? a comparative analysis Planta, 218 (3), 350-359 DOI: 10.1007/s00425-003-1113-5
Tamagnini, P., Leitao, E., Oliveira, P., Ferreira, D., Pinto, F., Harris, D., Heidorn, T., & Lindblad, P. (2007). Cyanobacterial hydrogenases: diversity, regulation and applications FEMS Microbiology Reviews, 31 (6), 692-720 DOI: 10.1111/j.1574-6976.2007.00085.x
Tenenbaum, D.J. (2008) Food vs. Fuel: Diversion of Crops Could Cause More Hunger. Environmental Health Perspectives 116: A224-A257.
Tikhonovich, I., & Provorov, N. (2007). Cooperation of plants and microorganisms: Getting closer to the genetic construction of sustainable agro-systems Biotechnology Journal, 2 (7), 833-848 DOI: 10.1002/biot.200700014
Towe, K. (1996). Environmental oxygen conditions during the origin and early evolution of life Advances in Space Research, 18 (12), 7-15 DOI: 10.1016/0273-1177(96)00022-1
Tsygankov, A. (2007). Nitrogen-fixing cyanobacteria: A review Applied Biochemistry and Microbiology, 43 (3), 250-259 DOI: 10.1134/S0003683807030040
WILLE, M., KRAMERS, J., NAGLER, T., BEUKES, N., SCHRODER, S., MEISEL, T., LACASSIE, J., & VOEGELIN, A. (2007). Evidence for a gradual rise of oxygen between 2.6 and 2.5Ga from Mo isotopes and Re-PGE signatures in shales Geochimica et Cosmochimica Acta, 71 (10), 2417-2435 DOI: 10.1016/j.gca.2007.02.019




