

These organic analogs to popsicle sticks and rubber bands often occur in interwoven networks reminiscent of the geodesic domes that Buckminster Fuller promoted in the 1950’s. Since form follows function, it’s hardly surprising that these natural geodesics offer the same benefits in nature that they do in architecture: high strength, dynamic stability, and material frugality.
Buckminster Fuller loved portmanteau words. He referred to his domes’ structural strategy as “tensegrity”, the marriage of tension and structural integrity. We often associate structural integrity with built-up, heavy structures intended to stand up to compressive loads, such as the massive concrete pillars that support the historic Fern Bridge. Compare this to the spindly but no less stable design of the historic Folsom Truss bridge. It’s easier to bend a thin metal bar than a concrete column, but the design of a truss eliminates any bending, shearing, or twisting forces. This leaves thin metal members to do what they do best—resist forces that push and pull them in the direction of their length.

Left: The Fern Bridge, built in 1911 in the Eel River Valley, CA, uses concrete in a complex state of compression to maintain its stability. Image credit: sunnyfortuna.com. Right: The Folsom Truss Bridge, built in 1895 in Folsom, CA, uses lightweight members exclusively in simple tension and compression. Image credit: myfolsom.com.
The triangular network of the Folsom bridge is a common motif in both manmade and evolutionary engineering. The reason for this pervasiveness is simply that the triangle is a very stable structure. It’s a heck of a lot easier to build a house of cards out of triangles than rectangles. Geodesic structures, such as Bucky’s domes and the human spine, are a special set of trusses that create curved forms from straight line segments. Some, like the trabeculae of human bones, echo the rigid structure of a manmade truss in order to create lightweight stiffness. But if frequency is any indication, nature’s best use for geodesic forms is in applications where stiffness is undesirable.
In lieu of hard bones, soft invertebrates such as nematodes use a “hydrostatic” skeleton. Muscles and skin contain their largely fluidic innards at a high pressure, like a water balloon. But unlike a water balloon, nematodes change their shape. They are capable of feats of extension, narrowing, and locomotion far more complex than their musculature dictates.
Recently in The Journal of Experimental Biology, Robert Shadwick discussed Clark and Cowey’s 1958 study revealing the role of inextensible collagen fibers in nemerteans and turbullarians. Their musculature is complemented by a lattice-like helical network of fibers resembling the form of a Chinese finger trap in both form and function. If a wormlike creature’s muscles contract its diameter, this fiber lattice reacts to extend the worm’s length. This breakthrough inspired a breadth of new discoveries in zoological morphology. Similar fibrous networks are being found in a wide array of applications, including elephant trunks, fish skin, lizard tongues, and even the human penis.
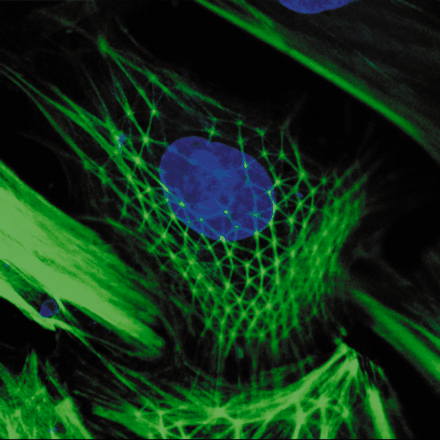 Cells are also squishy things in need of structure, and their cytoskeleton includes a tensile network of actin fibers that looks suspiciously like Disneyland’s Spaceship Earth dome. Researchers such as Donald Ingber of MIT are showing that this geodesic array does more than reinforce the cell’s shape. Ingber’s work focuses on the coupling of the cell’s force sensors (“mechanotransducers”) to its tensile fiber network.
Cells are also squishy things in need of structure, and their cytoskeleton includes a tensile network of actin fibers that looks suspiciously like Disneyland’s Spaceship Earth dome. Researchers such as Donald Ingber of MIT are showing that this geodesic array does more than reinforce the cell’s shape. Ingber’s work focuses on the coupling of the cell’s force sensors (“mechanotransducers”) to its tensile fiber network. When an event causes a cell to adhere to its surroundings, force changes in the cytoskeleton delivers a message to the cell. We are just beginning to understand that the cell reacts to mechanical signals like in dramatic ways; cellular forces dictate cell migration, instigate healing, and even trigger chromosomal events. Researchers are betting that understanding the structural underpinnings of bodily cells might lead to breakthroughs in the treatment of cancer, athsma, osteoporosis, wounds, and other human maladies.
Though it never caught fire as a housing concept, “tensegrity” is experiencing its second coming as a scientific buzzword. Buckminster Fuller might be cheered today to know that our every tissue represents a sprawling neighborhood of cellular geodesic dome-dwellers, showing nature’s endorsement of a great design strategy.
References:
Levin, S. The Tensegrity-Truss As A Model For Spine Mechanics: Biotensegrity. Journal of Mechanics in Medicine and Biology, 2003. 2: 375-388
Shadwick, RE. Foundations of Animal Hydraulics: Geodesic Fibres Control The Shape Of Soft Bodied Animals. J Exp Biol, 2008. 211: 289-291
Kier, W.M. and Smith, K.K. Tongues, tentacles and trunks: The biomechanics of movement in muscular-hydrostats. Zoological Journal of the Linnean Society , 1985. 83: 307-324.
Hebrank, MR. Mechanical properties and locomoter functions of eel skin. Biology Bulletin, 1980. 158:56-58.
Kelly, D. Penises as variable-volume hydrostatic skeletons. Annals of the New York Academy of Sciences, Reproductive Biomechanics, 2007. 1101(1), 453-463.
Ingber, DE. Tensegrity I. Cell structure and hierarchical systems biology. Journal of Cell Science, 2003. 116: 1157-1173 (2003).
Ingber DE. Tensegrity and mechanotransduction. Journal of Bodywork and Movement Therapies, 2008. 12(3) 198-200.



Comments