


Cell migration, which is involved in wound healing, cancer and tumor growth, and embryonic growth and development, has been a topic of interest to mathematicians and biologists for decades.
In a recent paper by Kristen Harley, Peter van Heijster, Robert Marangell, Graeme Pettet, and Martin Wechselberger, they write of a model describing cell invasion through directional outgrowth or movement in the context of malignant tumors, in particular, melanoma or skin cancer. Tumor cells move up a gradient, based on the presence of a chemical or chemoattractant – this process is called haptotaxis. Receptors on the exterior of cell walls detect and allow passing of the chemoattractant. Based on the locations of these receptors, cells determine the most favorable migration direction.
Continuum mathematical models that describe cell migration usually give rise to traveling waves—waves in which the medium moves in the direction of propagation at a constant speed. In this paper, the authors prove the existence and uniqueness of traveling waves to the model of malignant tumor invasion.
The model described takes into account the speed of the traveling waves, which corresponds to the rate of invasion of cells, as well as the extracellular matrix concentration (the medium surrounding cells that provides structural and biochemical support to cells). Movement of cells through diffusion is omitted as it is shown to play a relatively small role in the migration process.
Mathematically modeling species dispersal
Dispersal is an ecological process involving the movement of an organism or multiple organisms away from their birth site to another location or population where they settle and reproduce. An important topic in ecology and evolutionary biology, dispersal can either be random or directed. Random movement, as the name indicates, describes dispersal patterns that are unbiased and random, whereas directed or biased movement occurs when organisms sense and respond to local environmental cues by moving directionally. Dispersal is dependent on a variety of factors such as climate, food, and predators, and is often biased.
Fitness-dependent dispersal is a type of biased dispersal; the fitness of a species is given by its per capita growth rate. In many mathematical models of fitness-dependent dispersal, movement of organisms into and out of an area or region depends on the fitness differences between the organisms' resident patch and other patches in the habitat, and there is a net movement from patches of lower to higher fitness.
In a recent paper published in the SIAM Journal on Mathematical Analysis, authors Yuan Lou, Youshan Tao, and Michael Winkler propose a continuous-time and continuous-space reaction diffusion model for fitness-dependent dispersal where the species moves upward along its fitness gradient.
In ecology, ideal free distribution refers to the way in which organisms distribute themselves among patches proportional to the amount of resources available in each patch. Such a distribution minimizes resource competition and maximizes fitness. Thus it is natural to expect that dispersal strategies leading to ideal free distribution of populations would be favored over the course of evolution. The authors, in this paper, determine that fitness-dependent dispersal conveys advantages to approaching such ideal free distribution.
A model for Bluetongue disease dynamics in cattle
In a paper recently published in the SIAM Journal on Mathematical Analysis, authors Stephen Gourley, Gergely Röst, and Horst Thieme model disease persistence of a virus called Bluetongue using a system of several delay differential equations. The disease affects sheep and cattle, and is spread by biting midges. In sheep, the bluetongue virus can cause abortion, congenital abnormalities and death, though mild cases completely recover. In cattle, bluetongue does not generally cause death.
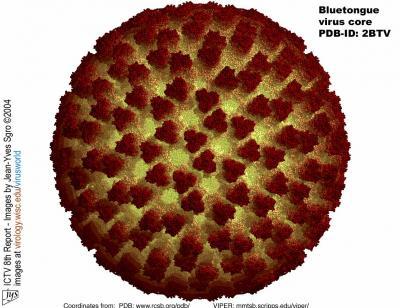
The atomic structure of the bluetongue virus core is shown. Ramsol image of PDB 2BTV by Dr. J.-Y. Sgro, UW-Madison. (Credit: Dr. J.-Y. Sgro, UW-Madison
The basic reproduction number for a disease is defined as the expected number of secondary cases produced by a single infection in a susceptible population. As in many infectious disease models, uniform disease persistence of bluetongue occurs if the basic reproduction number for the whole system exceeds one.
But an additional factor influences the disease state in the case of this disease, which is that it affects sheep much more severely than cattle. As a result, uniform disease persistence can occur in two different scenarios. If the disease reproduction number for the cattle-midge-bluetongue system with or without sheep is greater than one, bluetongue persists in cattle and midges even though it may eradicate the sheep, relying on cattle as a reservoir.
In the second situation, where the reproduction number of all host and vector species coexisting is greater than one, while the reproduction number for the cattle-midge-bluetongue system (without sheep) is less than one, bluetongue and all host and vector species coexist, and bluetongue does not eradicate sheep because it cannot persist on midges and cattle alone. The authors use different approaches of dynamical systems persistence theory to analyze the two situations.
References:
Approaching the Ideal Free Distribution in Two-Species Competition Models with Fitness-Dependent Dispersal
http://epubs.siam.org/doi/abs/10.1137/130934246
SIAM Journal on Mathematical Analysis, 46 (2), 1228-1262
Existence of Traveling Wave Solutions for a Model of Tumor Invasion http://epubs.siam.org/doi/abs/10.1137/130923129
SIAM Journal on Applied Dynamical Systems, 13 (1), 366-396
Uniform Persistence in a Model for Bluetongue Dynamics http://epubs.siam.org/doi/abs/10.1137/120878197 SIAM Journal on Mathematical Analysis, 46 (2), 1160-1184



Comments